by Michael Keenan
Ancient DNA (aDNA) was first extracted in 1984, by Russell Huguich and colleagues. The DNA was extracted from a 140 year old quagga tissue. This DNA was sequenced and was found to be mitochondrial in origin. Further success was achieved the following year by Svante Pääbo with the extraction of nuclear DNA from a 2,400 year old Egyptian mummy. These successes were in the 'pre-PCR' genomic era, but it was only after the development of PCR and its applications became apparent, that the study of ancient genetic material really gained the scientific community’s attention. Using PCR, combined with other established molecular techniques allowed many new experimental avenues to be explored. The common delimiting factor in these studies was the poor preservation of genetic material and/or its sequence integrity.
Ancient DNA is commonly found to be highly fragmented and composed of modified nucleotides. These features of aDNA come about as a result of the taphonomic processes following host death. Endogenous nucleases can retain their cleavage functionality for a variable period after death. This can give rise to the initial fragmentary genomic state. On a larger time scale, the continuing effects of processes such as, nucleotide hydrolysis, oxidation, deamination and depurination can cause substantial loss of sequence integrity. The rate and overall degree of this degradation is highly dependent on the particular system in which it occurs. Variables such as temperature, soil pH, DNA base composition and the particular complement of microorganisms present in the environment, all contribute to the 'end-state' of the host genome.
Frequently, in the case of aDNA studies, the tissue availability from which to extract DNA is limited. Since teeth and bones suffer degradation at a much lower rate than soft tissues, they are the final reservoirs of intact aDNA in many cases. This has led to teeth and bones becoming the main sources of aDNA used in archaeogenetic research and rigorous protocols are well established for both.
Ancient DNA (aDNA) was first extracted in 1984, by Russell Huguich and colleagues. The DNA was extracted from a 140 year old quagga tissue. This DNA was sequenced and was found to be mitochondrial in origin. Further success was achieved the following year by Svante Pääbo with the extraction of nuclear DNA from a 2,400 year old Egyptian mummy. These successes were in the 'pre-PCR' genomic era, but it was only after the development of PCR and its applications became apparent, that the study of ancient genetic material really gained the scientific community’s attention. Using PCR, combined with other established molecular techniques allowed many new experimental avenues to be explored. The common delimiting factor in these studies was the poor preservation of genetic material and/or its sequence integrity.
Ancient DNA is commonly found to be highly fragmented and composed of modified nucleotides. These features of aDNA come about as a result of the taphonomic processes following host death. Endogenous nucleases can retain their cleavage functionality for a variable period after death. This can give rise to the initial fragmentary genomic state. On a larger time scale, the continuing effects of processes such as, nucleotide hydrolysis, oxidation, deamination and depurination can cause substantial loss of sequence integrity. The rate and overall degree of this degradation is highly dependent on the particular system in which it occurs. Variables such as temperature, soil pH, DNA base composition and the particular complement of microorganisms present in the environment, all contribute to the 'end-state' of the host genome.
Frequently, in the case of aDNA studies, the tissue availability from which to extract DNA is limited. Since teeth and bones suffer degradation at a much lower rate than soft tissues, they are the final reservoirs of intact aDNA in many cases. This has led to teeth and bones becoming the main sources of aDNA used in archaeogenetic research and rigorous protocols are well established for both.
Applications
Identification of relationships:
The city of Pompeii was a Roman town in the Campania region of now southern Italy. In mid august 79 C.E Mount Vesuvius erupted; this was a catastrophic event which buried the town in 4 - 6m of sediment. The site was uncovered well over a thousand years later in 1748 by a team of excavators under the command of Rocque Joaquin de Alcubierre. An estimated ~1700 human remains are thought to be deposited at Pompeii, with over 1000 already excavated. Skeletal remains of 13 individuals were found in two adjacent rooms in the house known as Casa di Julius Polybius (house of Julius Polybius). Initial anthropological studies of the remains suggested 3 adult males, 4 boys, 3 adult females, 1 girl, as well as a child and unborn foetus of unknown sex. Two of the individuals showed evidence of spina bifida. The authors suggested possible familial relationships, indeed, two individuals were holding hands as is often seen in kin burials. More recently, work has been undertaken to try and genetically determine any possible relationships between these individuals as well as to confirm or refute the previous anthropological sex determination. Sex determination was carried out by amplification of the nuclear amelogenin locus and the genealogical aspect was tackled via mitochondrial DNA mtDNA sequence analysis. Since the X chromosome is common to both sexes and the Y is not; it is thought that reliability of male sex assignment is less than that of female sex assignment . Therefore, the team also amplified the alphiod locus, which is Y chromosome specific. Using these analyses, the team found there to be 8 males and 4 females. The unborn foetus could not be sexed. These results were in perfect support of the anthropomorphic data, with the added information that the previously unknown child was in fact a male. DNA was extracted from the left femur; competitive PCR and qPCR were used to quantify the amount of endogenous DNA molecules. Samples were then amplified for mtDNA regions spanning both hypervariable regions as well as a section of the coding region. Subsequent sequencing of products and analysis was carried out; these results are summarised in table 1 and the most informative data are shown schematically in figure 1.
The city of Pompeii was a Roman town in the Campania region of now southern Italy. In mid august 79 C.E Mount Vesuvius erupted; this was a catastrophic event which buried the town in 4 - 6m of sediment. The site was uncovered well over a thousand years later in 1748 by a team of excavators under the command of Rocque Joaquin de Alcubierre. An estimated ~1700 human remains are thought to be deposited at Pompeii, with over 1000 already excavated. Skeletal remains of 13 individuals were found in two adjacent rooms in the house known as Casa di Julius Polybius (house of Julius Polybius). Initial anthropological studies of the remains suggested 3 adult males, 4 boys, 3 adult females, 1 girl, as well as a child and unborn foetus of unknown sex. Two of the individuals showed evidence of spina bifida. The authors suggested possible familial relationships, indeed, two individuals were holding hands as is often seen in kin burials. More recently, work has been undertaken to try and genetically determine any possible relationships between these individuals as well as to confirm or refute the previous anthropological sex determination. Sex determination was carried out by amplification of the nuclear amelogenin locus and the genealogical aspect was tackled via mitochondrial DNA mtDNA sequence analysis. Since the X chromosome is common to both sexes and the Y is not; it is thought that reliability of male sex assignment is less than that of female sex assignment . Therefore, the team also amplified the alphiod locus, which is Y chromosome specific. Using these analyses, the team found there to be 8 males and 4 females. The unborn foetus could not be sexed. These results were in perfect support of the anthropomorphic data, with the added information that the previously unknown child was in fact a male. DNA was extracted from the left femur; competitive PCR and qPCR were used to quantify the amount of endogenous DNA molecules. Samples were then amplified for mtDNA regions spanning both hypervariable regions as well as a section of the coding region. Subsequent sequencing of products and analysis was carried out; these results are summarised in table 1 and the most informative data are shown schematically in figure 1.
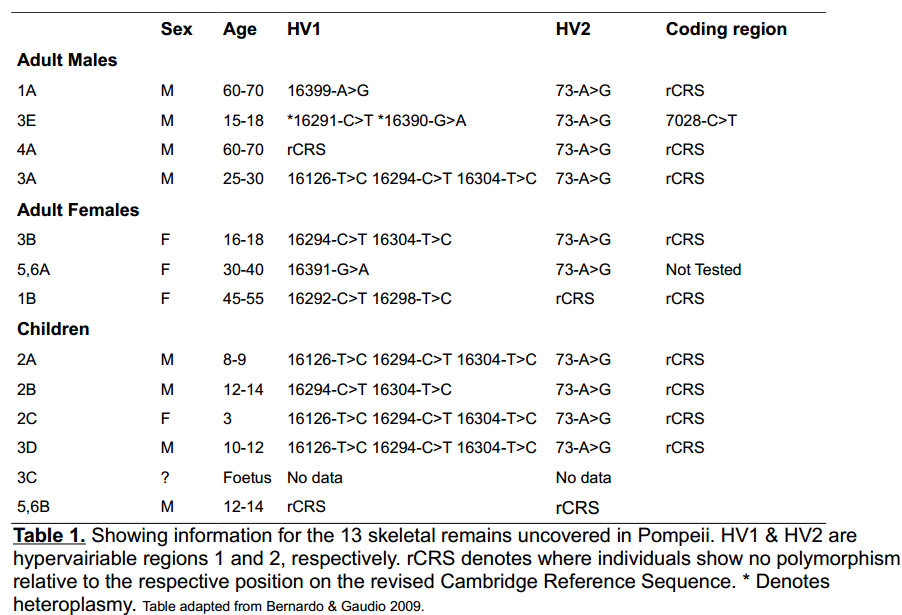
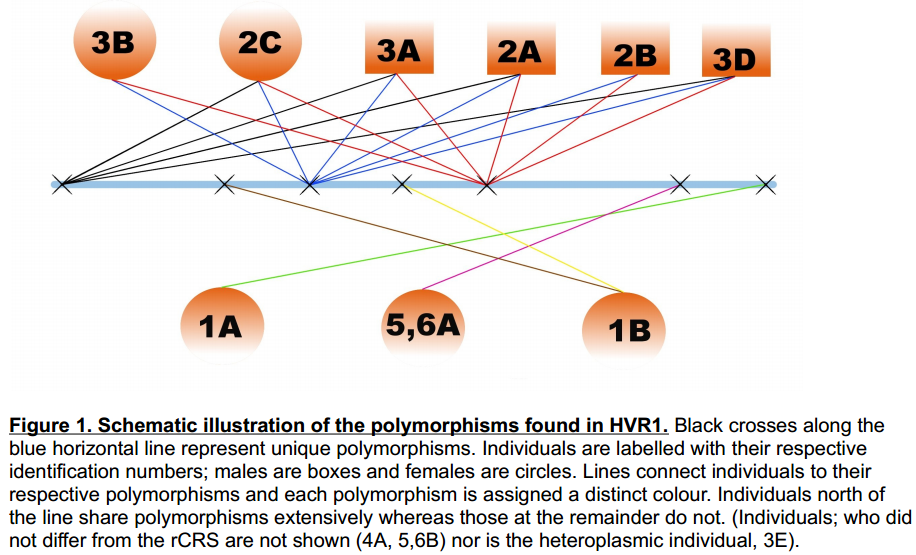
Individuals; 3B, 2C, 3A, 2A ,2B ,3D were determined to be of haplogroup T2b. The authors noted that
haplogroup T2b is the group assigned to 4.4 - 4.7% of modern Italians. They concluded that the probability of this haplogroup to occur in 50% of samples compared to the general level in the population was low enough to provide a strong indication of maternal ancestry. Females 2A and 3B both showed clear signs of spina bifida, further strengthening the case for familial relationships. Finally, a possible family network was laid out. This was based on the most likely scenario given the ages of the skeletons. 2C, 2A, 2B & 3D were suggested to be either siblings or cousins. 3B could be either the older sister, cousin or a maternal aunt of the four children. 3A is a male who could either be the father or a maternal uncle to the other 5 individuals.
This study shows how molecular evidence can contribute vital information to archaeological sites following excavation. For instance, traditional approaches toward sex determination primarily relied on the utilisation of morphological data. Unfortunately, as is the case with poorly preserved specimens, the telling features from which to base sex assignment can often be malformed or otherwise non-existent. And reliable familial grouping of specimens was near-impossible. Molecular analysis can sufficiently bypass these once-inescapable problems, even with very limited material.
haplogroup T2b is the group assigned to 4.4 - 4.7% of modern Italians. They concluded that the probability of this haplogroup to occur in 50% of samples compared to the general level in the population was low enough to provide a strong indication of maternal ancestry. Females 2A and 3B both showed clear signs of spina bifida, further strengthening the case for familial relationships. Finally, a possible family network was laid out. This was based on the most likely scenario given the ages of the skeletons. 2C, 2A, 2B & 3D were suggested to be either siblings or cousins. 3B could be either the older sister, cousin or a maternal aunt of the four children. 3A is a male who could either be the father or a maternal uncle to the other 5 individuals.
This study shows how molecular evidence can contribute vital information to archaeological sites following excavation. For instance, traditional approaches toward sex determination primarily relied on the utilisation of morphological data. Unfortunately, as is the case with poorly preserved specimens, the telling features from which to base sex assignment can often be malformed or otherwise non-existent. And reliable familial grouping of specimens was near-impossible. Molecular analysis can sufficiently bypass these once-inescapable problems, even with very limited material.

Identification of individuals:
The infamous Jesse James was a notorious American outlaw from Missouri; he was a member of the James-Younger Gang. In 1882, Jesse was shot in the back of the head whilst he was cleaning a dusty picture in his home. The killer was Robert Fort (one of the few people Jesse trusted at the time). Rumors of Jesse's survival emerged soon after his death; some even claiming he went on to father two children and lived under the name of 'Thomas Howard'. The presumptive remains of Jesse James were buried at the family residence in 1882 and subsequently moved to another plot in 1902.
In a paper published by Anne Stone and colleagues the remains were exhumed; hair, teeth and bone samples were collected. Samples were collected from both the initial burial site and the Mt. Olive site, and mtDNA was extracted. Hot-start PCR was employed to successfully amplify the hypervariable 1 region of the mitochondrial genome and the products were then sequenced. The gained sequence was then compared against the same genomic location for two of Jesse's maternal relatives. Individual 'RJ' was the living grandson of Jesse's sister Susan, whereas 'MN' was the living great-granddaughter of Susan. Both individuals share the same maternal lineage as Jesse James. The results are summarised in figure 2. As shown, all samples taken from the presumptive remains of Jesse James (C, F, H-1 & H-2) contain a single nucleotide polymorphism at 5 distinct positions (with respect to the Cambridge Reference Sequence). These sequences are derived from mtDNA which was sampled from both places of burial which suggests the DNA is genuine. Each SNP is common to both RJ and MN, suggesting that the remains are, indeed, those of Jesse James.
The infamous Jesse James was a notorious American outlaw from Missouri; he was a member of the James-Younger Gang. In 1882, Jesse was shot in the back of the head whilst he was cleaning a dusty picture in his home. The killer was Robert Fort (one of the few people Jesse trusted at the time). Rumors of Jesse's survival emerged soon after his death; some even claiming he went on to father two children and lived under the name of 'Thomas Howard'. The presumptive remains of Jesse James were buried at the family residence in 1882 and subsequently moved to another plot in 1902.
In a paper published by Anne Stone and colleagues the remains were exhumed; hair, teeth and bone samples were collected. Samples were collected from both the initial burial site and the Mt. Olive site, and mtDNA was extracted. Hot-start PCR was employed to successfully amplify the hypervariable 1 region of the mitochondrial genome and the products were then sequenced. The gained sequence was then compared against the same genomic location for two of Jesse's maternal relatives. Individual 'RJ' was the living grandson of Jesse's sister Susan, whereas 'MN' was the living great-granddaughter of Susan. Both individuals share the same maternal lineage as Jesse James. The results are summarised in figure 2. As shown, all samples taken from the presumptive remains of Jesse James (C, F, H-1 & H-2) contain a single nucleotide polymorphism at 5 distinct positions (with respect to the Cambridge Reference Sequence). These sequences are derived from mtDNA which was sampled from both places of burial which suggests the DNA is genuine. Each SNP is common to both RJ and MN, suggesting that the remains are, indeed, those of Jesse James.
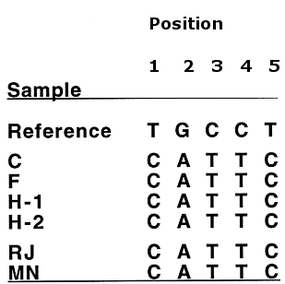
Figure 2. Showing single nucleotide polymorphisms (SNP) relative to the reference sequence. C, F, H-1, H-2 correspond to mitochondrial sequence extracted from Jesse James. RJ and MN correspond to maternal relatives of Jesse. All SNPs are common to all samples. (Position numbers are arbitrary.)
Authentication
Working with aDNA can carry a lot of potential pitfalls that the researcher must be aware of. The tried and tested genetic approaches such as DNA extraction and PCR amplification must be used within the context of aDNA research. PCR success rate and the final composition of PCR product is a highly variable factor in aDNA research. Large inconsistencies between pre-PCR sample composition, even between samples from a common burial site, can contribute to this variability. This should not be a surprise, since the processes occurring post-mortem are largely inextricable. It would be an erroneous conclusion to assume a shared burial site equates to a shared taphonomy. Ancient samples can contain inhibitory substances; these substances frequently co-extract with DNA and can present false negative results by interfering with efficient and reliable PCR amplification. Removal of these inhibitory substances is achieved by various experimental manipulations e.g. addition of bovine serum albumin, and results in a higher quality PCR product. Other manipulations may also be used to increase product quality such as hot-start PCR and competitive-PCR. Even with optimal extraction conditions, prudence suggests that any given sample from a past individual will be a mixture of endogenous and exogenous genetic material. If highly specific PCR primers are used then we may be reasonably confident that the PCR product is from human derived sequence. This however, does nothing to distinguish authentic aDNA and its modern counterpart. For instance, it is well known that PCR amplification is error prone. Taq, the DNA polymerase enzyme most commonly used in PCR, can introduce errors as it copies the template strand. A Taq induced mutation is indistinguishable from a genuine mutation and this may hamper haplotype assignment. Due to the exponential nature of PCR, if a non-authentic mutation occurs early in the reaction, then the final population of amplicons may contain a large abundance of these false polymorphisms. Further, Taq is not indiscriminate in terms of which fragments it amplifies, certain fragments may be amplified more efficiently than others whether they are the sequence we are interested in or not. Furthermore, mutational events which occur in the primer regions of authentic aDNA may render their respective polynucleotides unsuitable for PCR amplification.Since the starting material for an aDNA study is from the same species of the experimenter, there is no one way to occlude modern DNA contamination. Approaches to minimise its occurrence and identify its source are the only real effective strategies as of yet, (e.g. genotyping any individuals who have been in contact with the specimen). Molecular cloning has often been (perhaps ostensibly regarded as a vital stage in the pipeline from aDNA extraction to sequencing. Although it is completely possible to directly sequence PCR product, this is not the common approach for aDNA studies. There main rationale behind cloning comes from the idea that the PCR product may well be a population of differing fragments. Molecular cloning allows separation of this population so that each fragment may be analysed on its own. depending on the sequencing technology, loss of sequence information can occur at the ends of the sequence. If this is a putative aDNA fragment, then we may loose already limited and valuable sequence information. In the case of cloned PCR product; sequencing is initiated from vector sequence. This addition of vector sequence allows this end degradation problem to be largely bypassed as it will be restricted to the vector sequence. Furthermore, efficient PCR primers can be poorly suited as as sequencing primers, whereas very effective sequencing primers have been designed for most commonly used vectors. If sequence results do not make sense, or haplotype assignment is inconspicuous, then it is still a viable option to go back and sequence more clones to build a consensus sequence or identify any possible sources of the ambiguity (e.g. contamination, heteroplasmy). The use of cloning does indeed overcome many of the technical obstacles put forward here. It does not however, address the genuineness of whether PCR products to be cloned are authentic or otherwise. These questions have not yet been answered to an appropriate degree, so again, precautionary and preventative measures must be followed. It seems the most invaluable tool in ancient DNA study thus far, is the researcher's scrupulosity.
References
Stone, AC., Starrs, JE., Stoneking, M. (2001). Mitochondrial DNA analysis of the presumptive remains of Jesse James. Journal of Forensic Science. 46 (1), 173-176.
Bernardo, GD., Gaudio, SD., Galderisi, U., Cascino, A., Cipollaro, M.,. (2009). Ancient DNA and Family Relationships in a Pompeian House.Annals of Human Genetics. 73 (4), 429-437.
Bernardo, GD., Gaudio, SD., Galderisi, U., Cascino, A., Cipollaro, M.,. (2009). Ancient DNA and Family Relationships in a Pompeian House.Annals of Human Genetics. 73 (4), 429-437.
 RSS Feed
RSS Feed
